Un arbre est constitué de tissus vivants, qui forment un manchon interne relativement fin regroupant les fonctions essentielles (conduction de la sève, mise en réserve), et de tissus morts qui forment un
cylindre central et un manchon externe, que nous appelons communément l'écorce.
Chacun des différents tissus remplit un rôle.
 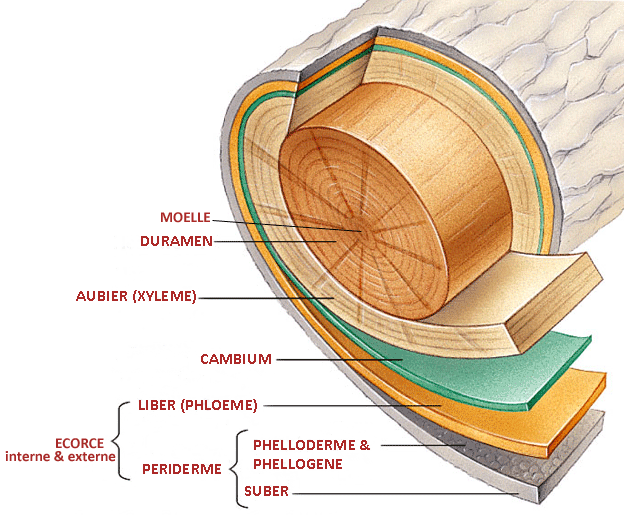
De l'intérieur vers l'extérieur, on trouve successivement:
- le DURAMEN
Le duramen ou "bois parfait" correspond aux zones d'accroissement les plus anciennes : c'est le coeur de l'arbre, la partie la plus dure et la plus résistante. Compact, dense et imputrescible, il ne comporte plus de cellules
vivantes et a un rôle de soutien, un peu comme un squelette.
Il entoure la moelle, un tissu végétal un peu mou qui se forme au début de la croissance, avant que le bois n'apparaisse.
- l'AUBIER (XYLEME SECONDAIRE)
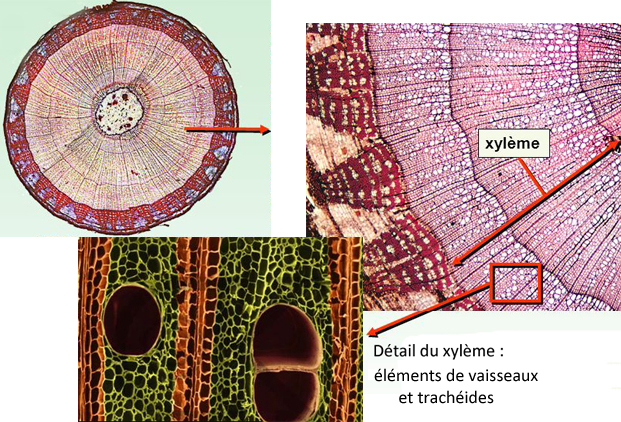
Le xylème (du grec ξυλον, bois) secondaire, communément appelé "bois" (ou "bois vivant") puisqu'il est lignifié, correspond aux zones d'accroissement
récentes. C'est un bois tendre, physiologiquement actif, qui fait donc partie des parties vivantes de l'arbre. Il est en général plus clair que le bois parfait, le plus souvent blanchâtre.
Le xylème est un tissu complexe tant sur le plan structural que fonctionnel ; il est constitué de :
-fibres qui assurent le soutien (chez les feuillus)
-cellules de remplissage ( parenchyme), dont la fonction est la mise en réserve ;
-vaisseaux ligneux de deux sortes spécialisés dans la conduction de la sève brute, qui est le rôle principal du xylème secondaire.
Ces vaisseaux sont formés de cellules mortes (plus précisément : de cellules qui se sont sclérifiées et vidées de leur contenu), alignées les unes par rapport aux autres :
-chez les gymnospermes (=conifères), ces vaisseaux sont uniquement des trachéides, composés de cellules étroites et allongées, superposées,
terminées en biseaux, ce qui provoque une circulation en chicane, à travers des perforations appelées ponctuations ;
-les angiospermes (=feuillus) possèdent principalement des éléments de vaisseaux, constitués de cellules larges et courtes, disposées bout à
bout, dont les parois longitudinales se sont épaissies par des dépôts de lignine et dont les parois transversales ont disparu, créant ainsi de véritables tubes, où se fait verticalement la circulation.
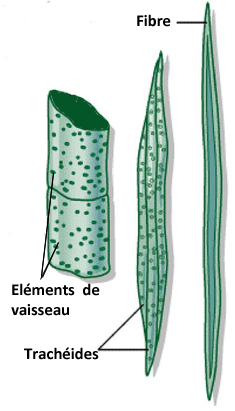 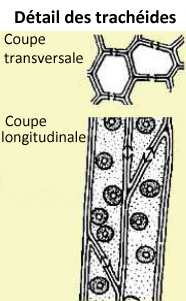 
Ces vaisseaux communiquent latéralement entre eux pour permettre à la sève de continuer à circuler si l'un d'eux vient à s'obstruer.
Périodiquement, les vaisseaux situés le plus à l'intérieur se bouchent et cessent
d'alimenter l'arbre : ils se transforment alors en duramen.
Chaque année, une nouvelle couche de xylème se forme, produisant un cerne : le nombre total de cernes indique l'âge de l'arbre.
- le CAMBIUM
Le cambium (du latin cambiare changer) est un feuillet très fin, de couleur verte, constitué de cellules méristématiques,
c'est à dire de cellules indifférenciées formant une zone de croissance : ces cellules ont la faculté de se diviser activement puis de se différencier en acquérant une structure et une fonction particulières, pour assurer le
développement de la plante ; autrement dit, elles peuvent produire n'importe quelle cellule spécialisée (cellules de vaisseaux, cellules de parenchyme etc)
Le cambium, qui assure le grossissement de l'arbre (=sa croissance en diamètre) est un méristème secondaire : il produit du bois (xylème secondaire) vers l'intérieur et du liber (phloème
secondaire) vers l'extérieur, mais il forme beaucoup moins de liber que de bois.
Il peut aussi, en cas de blessure, former un bourrelet cicatriciel dont les bords vont finir pas se rejoindre ; il peut également générer de
nouvelles racines -faculté que l'on utilise dans le bouturage ou le marcottage- ou souder un greffon, en réalisant une connexion vasculaire pour l'alimenter.
- le LIBER (PHLOEME SECONDAIRE )
Le phloème secondaire constitue l'écorce interne. C'est un tissu complexe, qui comporte des:
-cellules de remplissage ( parenchyme), dont la fonction est la mise en réserve ;
-éléments conducteurs, dévolus à la distribution de la sève élaborée entre les sources (tissus photosynthétiques ou cellules de réserves) et toutes les régions de l'arbre (=
transport multidirectionnel).
Ces éléments conducteurs sont constitués de :
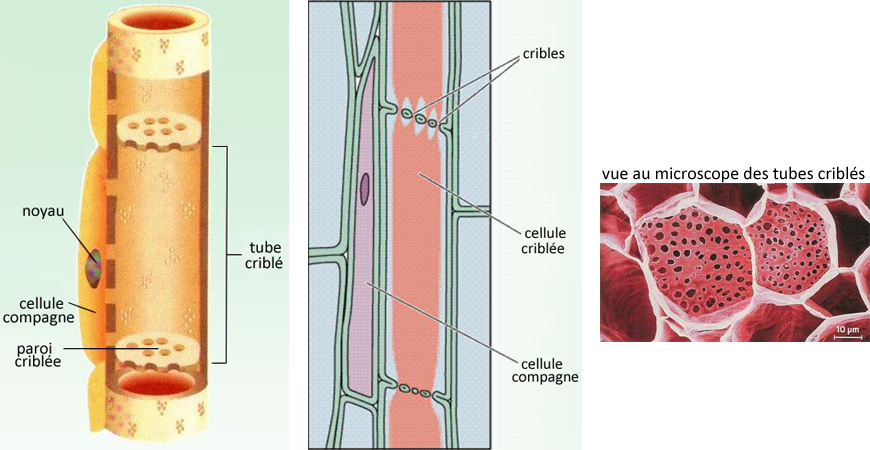
-tubes criblés, formés de cellules disposées en files verticales ; ces cellules n'ont ni noyau ni vacuole, mais sont vivantes, contrairement à
celles du xylème ;
-cellules compagnes, qui bordent les tubes criblés et assurent le transfert des sucres vers les cellules criblées ou leur déchargement dans les
cellules-cibles traversées par le système conducteur.
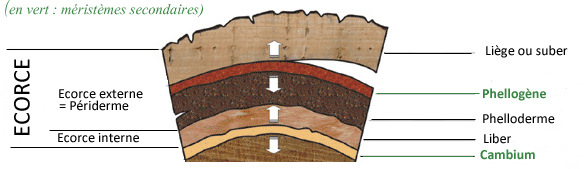
- l'ECORCE EXTERNE ou PERIDERME
Le périderme est une enveloppe protectrice efficace contre la déshydratation, le gel, la pluie, les rayons du soleil, les variations de température, les insectes etc ; il se met en place à la disparition de l'épiderme
(premier tissu de protection, superficiel, constitué d'une seule couche de cellules vivantes jointives), lors de la formation des méristèmes secondaires.
En effet, c'est le deuxième méristème secondaire, appelé phellogène, qui crée les deux autres tissus du périderme. Il produit
-vers l'intérieur une très fine couche de cellules vivantes, appelée le phelloderme, tissu parenchymateux à fonction d'accumulation de réserves
essentiellement ;
-vers l'extérieur des cellules mortes très jointives, remplies d'air, qui forment le liège ou suber.
Le liège est imprégné de subérine, une macromolécule à consistance cireuse, qui le rend imperméable autant à l'air qu'à l'eau ; il peut également renfermer des tanins ou des résines
assurant un rôle protecteur contre les attaques d'organismes pathogènes.
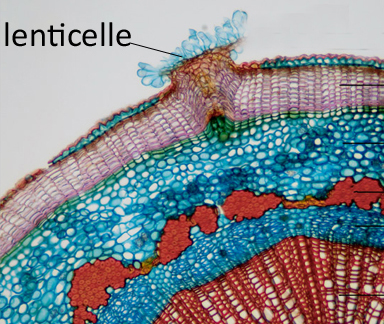
Il est déchiré par endroits par des lenticelles, petites taches poreuses formant des aspérités, qui permettent les échanges gazeux entre l'atmosphère et les tissus internes.
Au bout d'un certain nombre d'années, un ou plusieurs phellogènes -discontinus et se chevauchant plus ou moins entre eux- se forment sous le phellogène initial et prennent son relais : toutes les cellules vivantes extérieures
au phellogène le plus interne sont alors vouées à mourir.
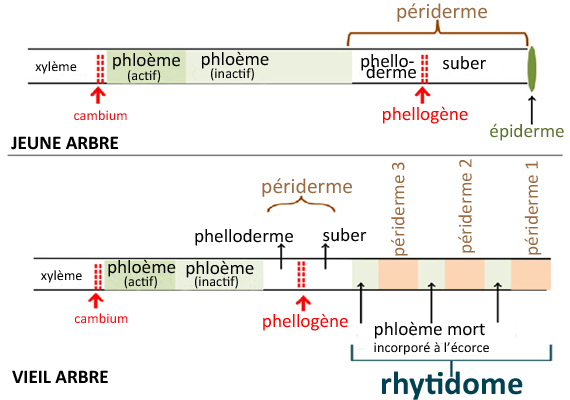
Les nouveaux phellogènes incorporent vers l'intérieur le phloème de l'année, écrasé par la croissance, et progressent vers l'extérieur, en recoupant ou en éliminant les anciens phellogènes situés au-dessus, en incorporant
vers l'extérieur les anciens phloèmes ainsi que les phellodermes et le suber des anciens péridermes.
C'est ce qui donne naissance au rhytidome -ce que nous appelons communément
"écorce"- caractérisé par sa surface fragmentée et/ou sa rugosité.
Quelques arbres cependant ont des phellogènes de forme annulaire, qui produisent une écorce quasiment lisse.
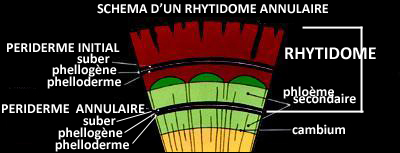 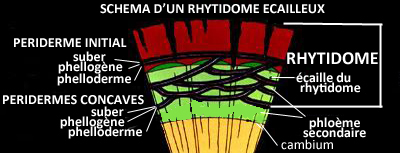
L'écorce se desquame généralement sous la pression des tissus sous-jacents, par plaques, écailles, bandes ou anneaux complets, sauf chez certaines espèces chez lesquelles elle s'épaissit au fur et à mesure avant, éventuellement,
de tomber par morceaux (chêne, séquoia...).
   |
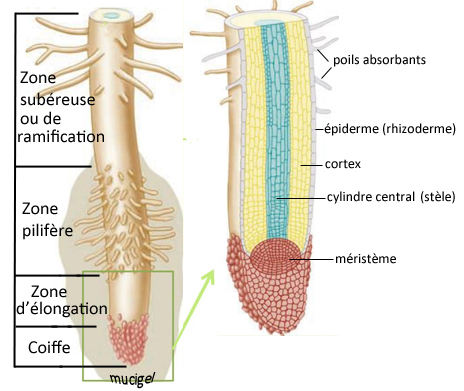 
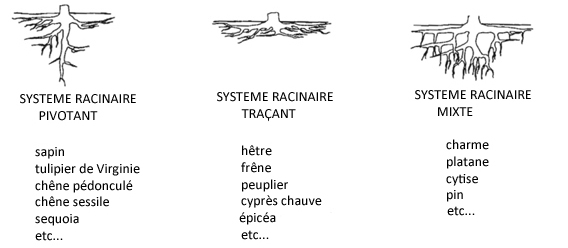
Dans le sens longitudinal, les racines comportent quatre parties :
- la zone subéreuse, partie dépourvue de poils, où se développent les ramifications ;
- la zone pilifère (du latin pilum poil et fero porter), qui émet les
poils absorbants, prolongements filamenteux de quelques millimètres de longueur (pour un diamètre de 12 à 15 micromètres) offrant à la plante une énorme surface de contact avec son milieu ;
- la zone d'élongation ou d'allongement, partie sans poils ni ramifications, abritant à son extrémité le méristème apical racinaire où s'opèrent les divisions cellulaires qui permettent la
croissance en longueur de la racine ;
- la coiffe, enveloppe terminale dont les cellules se renouvellent constamment, assurant la protection du méristème et permettant la pénétration de la racine dans le sol.
Une grande partie de la racine est enrobée d'une substance visqueuse, qu'elle produit elle-même, appelée mucigel.
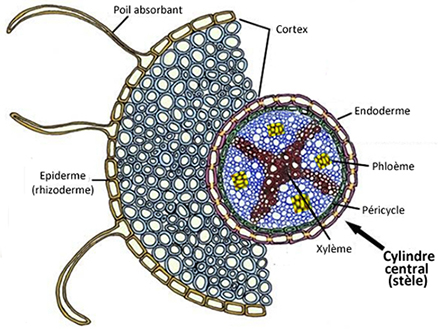
Transversalement, les racines comportent deux parties :
- l'écorce, comprenant :
-le rhizoderme (=épiderme), pilifère ou subérifié ;
-le cortex (ou plus précisément parenchyme cortical), situé sous l'épiderme et constitué de cellules irrégulières à grandes
vacuoles pour stocker les réserves ;
-l'endoderme, assise la plus interne de l'écorce, qui sépare le cortex du cylindre central ; il est constitué d'une couche unique de cellules jointives, dont la
perméabilité permet néanmoins aux molécules d'eau et d'oxygène de passer.
- le cylindre central ou stèle, situé au coeur de la racine, qui comporte :
-le péricycle, assise externe du cylindre central, faite d'une couche de cellules qui vient doubler l'endoderme et à partir de laquelle vont se former les
ramifications ;
-les faisceaux ligneux ( xylème) et libérien ( phloème), qui assurent le transport de la sève et qui se trouvent dans le
parenchyme médullaire.
Les méristèmes secondaires (cambium et phellogène), qui permettent la croissance en épaisseur des racines, se mettent en place selon un processus complexe :
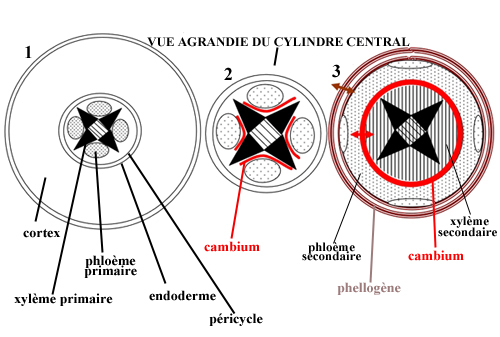 |
1. La jeune racine est dépourvue de tissus secondaires : elle ne comporte que le xylème primaire et le phloème primaire, au centre du péricycle.
2. L'installation du cambium se fait à partir de cellules du péricycle ; de forme sinusoïdale, il vient entourer le xylème primaire.
3. Le cambium prend une forme circulaire, il produit du bois (xylème secondaire) vers le centre et du liber (phloème secondaire) vers l'extérieur, en écrasant le phloème primaire.
Le phellogène, un deuxième méristème annulaire dont l'installation se fait elle aussi à partir du péricycle, produit l'écorce (suber) sur sa face externe et le phelloderme, très peu abondant, sur sa face interne.
Cette croissance à partir de l'intérieur a pour effet le déchirement, puis l'exfoliation des tissus les plus extérieurs (cortex, épiderme). |
|